

To see this report in Portable Document
Format (PDF), please click on the button. ![]() Should you not have Adobe
Acrobat Reader (required to read PDF files), this free program
is available for download at http://www.adobe.com/prodindex/acrobat/readstep.html
Should you not have Adobe
Acrobat Reader (required to read PDF files), this free program
is available for download at http://www.adobe.com/prodindex/acrobat/readstep.html
Open Forum: In an attempt to promote free and open discussion of issues, The Agrichemical and Environmental News encourages letters and articles with differing views. To discuss submission of an article, please contact Dr. Allan Felsot at 509-372-7365 or afelsot@tricity.wsu.edu; Dr. Catherine Daniels at 509-372-7495 or cdaniels@tricity.wsu.edu; or Dr. Doug Walsh at 509-786-2226 or dwalsh@tricity.wsu.edu. The newsletter is available in a hardcopy version for a $15 yearly subscription fee. Please contact newsletter editor Sally O'Neal Coates at 509-372-7378 or scoates@tricity.wsu.edu for details.
This page has been accessed times since May 24, 2000.
In the February and March issues of Agrichemical and Environmental News (AENews Nos. 166 and 167), I shared some background on the concerns surrounding children's exposure to pesticides and I outlined University of Washington (UW) studies on children in the Wenatchee area. In the last week of April, a new analysis of our Wenatchee studies was published in Environmental Health Perspectives, a scientific journal sponsored by one of the National Institutes of Health (see Editor's Note at end of article). Once published, a paper like this can become news, and this one did. The information released by the University of Washington Office of Health Sciences and Medical Affairs was headlined: "UW Study Finds Many Farm Children Are Exposed to Pesticides." This was translated in the Seattle Times on April 25, 2000, as "Kids' Pesticide Levels Unsafe."
Why did our findings draw media attention? Did our paper really demonstrate that children are exposed to pesticides at "unsafe" levels?
In our report, we tried to answer the question that parents and others ask when they learn about our studies of children and pesticides: "What are the risks? Are the levels safe?" These are not easy questions to answer.
Our study evaluated the exposures of 109 children living in Chelan and Douglas counties. Most (91) had parents working in agriculture. The others (18) did not have any household members involved in agriculture, and lived at least one-quarter mile from treated farmland. The metabolites we measured in the children's urine are common to several organophosphorus (OP) pesticides, including azinphos-methyl and phosmet. Our approach was to convert the OP pesticide metabolites found in the urine of children to estimates of the total amount of pesticides the children probably absorbed on the day we sampled. These dose estimates were then compared to guidelines developed by the Environmental Protection Agency and the World Health Organization (Table 1).
|
|
||
| Children's OP pesticide doses relative to the U.S. Environmental Protection Agency chronic dietary reference doses (RfDs), and World Health Organization acceptable daily intakes (ADIs) for azinphos-methyl and phosmet1 | ||
| Regulatory Reference Value |
|
|
|
|
||
| EPA Chronic Reference Dose (RfD) | ||
| Azinphos-methyl (1.5 µg/kg/d) | 56 | 44 |
| Phosmet (11 µg/kg/d) | 8.9 | 0 |
| WHO Acceptable Daily Intake (ADI) | ||
| Azinphos-methyl (5 µg/kg/d) | 19 | 22 |
| Phosmet (20 µg/kg/d) | 3.3 | 0 |
| 1Includes all children in the study; assumes doses are attributable entirely to either azinphos-methyl or phosmet. | ||
| 2Based on 91 estimates for agricultural children and 18 estimates for reference children. | ||
Our analysis assumed that the metabolites were the result of exposure to either azinphos-methyl or phosmet, the two chemicals found in nearly all of the housedust samples we collected from the children's homes. The results are presented in Table 2, and a graph of dose distribution is presented as Figure 1.
|
|
||||
| Spray season dose estimates1 adjusted by daily creatinine output. Children were aged 0-6 years. Doses were based on two dialkylphosphate metabolites (DMTP and DMDTP) common to the dimethyl organophosphorus pesticides. | ||||
|
|
||||
|
|
|
|
|
|
| Median |
|
|
|
|
| 25th percentile |
|
|
|
|
| 75th percentile |
|
|
|
|
| Mean |
|
|
|
|
| Std. Dev. |
|
|
|
|
| Range |
|
|
|
|
| 1Spray season dose estimates were based on the average of two samples per child. Only one child was used from each family in this analysis. All samples were collected during the May-July spraying season. In cases with missing samples, a single sample was used to estimate average dose. | ||||
| 2Agricultural children are a combination of applicator and farmworker children. | ||||
| 3Applicator and farmworker children dose estimates were not statistically different (Mann-Whitney U Test). | ||||
| 4Applicator and reference children dose estimates were statistically different (p=0.05, Mann-Whitney U Test). | ||||
| 5Agricultural and reference children dose estimates were marginally different (p=0.06). | ||||
|
|
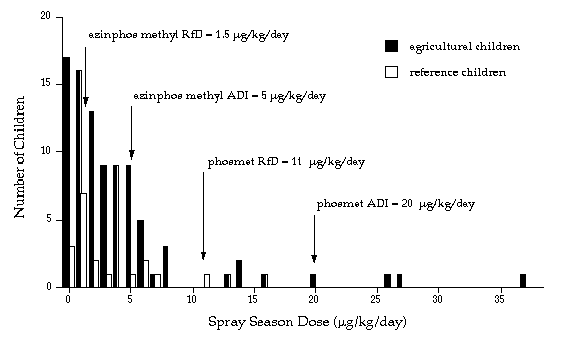 |
| Distribution of OP pesticide dose estimates for children in an agricultural community, derived from urinary metabolite measurements and adjusted for creatinine concentration. Spray season dose estimates for 109 children (91 agricultural, 18 reference). The doses are expressed as micrograms of pesticide per kilogram of body weight. Arrows indicate guidelines that have been established by EPA and WHO for azinphos-methyl and for phosmet. Azinphos-methyl is a more toxic OP pesticide than phosmet, so its EPA reference dose (RfD) and WHO acceptable daily intake (ADI) levels are lower. |
We found that for children whose parents worked in agriculture as either orchard applicators or field-workers, more than half of the doses estimated for the spray season exceeded the U.S. Environmental Protection Agency's chronic dietary reference dose (RfD) and about one fifth exceeded the World Health Organization's acceptable daily intake (ADI) values for azinphos-methyl. For children whose parents did not work in agriculture the values were 44% and 22%, respectively. When we considered that the metabolites were due to phosmet exposure, we found that less than 10% of the children exceeded the EPA and WHO reference values. None of the dose estimates exceeded what is called the "no effect" level determined in animal studies. We also noted that the study took place during a period of active spraying, and we cautioned readers not to generalize to other times of the year, or other regions. Nevertheless, it seems reasonable to assume that these children were exposed at these levels across the 40 to 50 days of the spraying season.
What do these numbers and comparisons really mean for children's health? The major public health message is that these findings are cause for concern, but not for alarm. We can say with some certainty that these exposures fall short of causing acute health effects, since the WHO and EPA guidelines incorporate large uncertainty factors. But it is also clear that the exposures for many of these children fall into that zone of uncertainty.
Some will argue that the current guidelines are too stringent, but others argue that they are not protective enough, particularly for children. Current regulatory methods are based on measurements of residues in food, water, and the environment, from which models are developed to estimate dose. Often these models include very conservative or protective assumptions, which can lead to high estimates and the appearance of risk that may or may not be present. Biological monitoring data are not normally used in the regulatory process, as they are very cumbersome to obtain and complex to coordinate. Yet it seems clear that they can provide a more accurate estimate of the dose that a child receives. The primary scientific message is that biological measurements, such as pesticide metabolites in urine, can give us reasonably good estimates of dose and risk. As we monitor more children we will be able to see patterns that can aid in developing commonsense and cost-effective methods to reduce exposures.
Dr. Richard Fenske is Professor of Environmental Health at the University of Washington's School of Public Health and Community Medicine, and Director of the Pacific Northwest Agricultural Safety and Health Center (PNASH). He also serves on EPA's Science Review Board, a congressionally mandated advisory board for pesticide science policy. He can be reached at rfenske@u.washington.edu or (206) 616-1958.
|
|
Return to Table of Contents for the June 2000 issue

The miticide/insecticide abamectin (Agrimek) was first used against two-spotted spider mite (Tetranychus urticae, TSSM) on Washington hops in 1991. Thus, as we now enter its tenth season of use, we thought it timely to re-check the susceptibility of hop yard mites to abamectin.
To date, hop growers have had little complaint about abamectin's performance, although there were some reports of reduced control during the hot 1998 season. Resistance to abamectin has been detected in spider mite populations on a number of other crops (e.g., apples, roses) in the United States and Europe (1,2).
In order to determine whether TSSM from hop yards exhibit a decreased level of susceptibility to abamectin, we needed to establish a baseline abamectin toxicity level for TSSM.
First, we collected TSSM from a Kennewick backyard. These specimens probably had never been exposed to abamectin. Next, we conducted tests on these susceptible mites using a Potter Precision Spray Tower, a device that delivers known quantities of pesticide very accurately and efficiently. Using a range of concentrations of abamectin, we were able to develop a dose-mortality line and determine the lethal concentration (LC) of chemical needed to kill 50 or 99% of the TSSM population. (These amounts are known as the LC50 and LC99, respectively.)
Finally, we performed identical tests on four TSSM strains obtained from three commercial hop yards and a hop yard at WSU's Irrigated Agriculture Research and Extension Center (IAREC) during 1999. Each TSSM strain was cultured separately on dwarf bean plants at WSU-IAREC and tests were replicated three or four times.
Our backyard TSSM strain was very susceptible
to abamectin, with a concentration of 0.1 parts per million (ppm)
killing 100% of individuals tested (i.e., LC99 of <0.1). The
LC50 was 0.01, meaning fifty percent of individuals died when
exposed to 0.01 ppm. These results were consistent with those
for other susceptible TSSM strains reported in the scientific
literature. In contrast, the four hop yard strains had LC99s of
2.5 to 5.0, indicating a resistance factor of 25- to 50-fold.
The differences between the LC50 values were even greater, suggesting
a resistance factor of 75- to 100-fold. In comparison, a previous
study of TSSM strains from apple and pear orchards in Washington
during 1994-95 showed resistance levels to abamectin of 5- to
27-fold (1). Campos et al. examined
TSSM strains from nurseries in California, Florida, the Canary
Islands and Holland and found abamectin resistance levels ranging
from 0.5- to 175-fold (2).
Given the relative consistency of our results between mites from the four hop yards, it is likely that hop yard TSSM in the Yakima Valley are indeed harder to kill than strains from abamectin-free locations.
So what does this mean to the hop grower, preparing to mount his annual mite control campaign? Judging from most grower reports, abamectin is still an effective product, providing good mite control in hop yards. At the current recommended application rate of 16 fluid ounces per acre, hop growers are treating mites with a concentration that is close to 100 ppm. So it is not surprising that abamectin-resistant mites, which require only 2.5 to 5.0 ppm for total kill, are still being successfully controlled. However, we may have reached a critical stage with this product where overuse could precipitate even higher levels of resistance and eventual control failure. Thus, if at all possible, we recommend that hop growers only use abamectin once each season to help suppress further resistance development. If re-treatment is necessary, a miticide from another chemical group should be used.
Abamectin has the potential to remain an effective and valuable miticide in hops for many more years. Correct and judicious use (i.e., once a season, applied at the full rate with complete and thorough spray coverage of the crop) should ensure maximum longevity of abamectin in Washington hop yards.
WSU will closely monitor abamectin resistance in hop yard TSSM during the coming seasons. We invite hop growers to alert us of difficulties in controlling mites with this product. This will allow us to determine whether control problems are caused by increased resistance or some other factor.
Dr. David James is an Assistant Entomologist with Washington State University's Irrigated Agriculture Research and Extension Center (IAREC) in Prosser. He can be reached at djames@tricity.wsu.edu or (509) 786-9280. Agricultural Research Technician Tanya Price's e-mail is tprice@tricity.wsu.edu.
Return
to Table of Contents for the June 2000 issue
No pest control technique--with the arguable exception of the flyswatter--is immune to the possibility of resistance. Over 400 insect pest species are estimated to have developed resistance to one or more classes of insecticides (3).
Of all the ecological concerns regarding the Bt toxin, resistance is the one that has already occurred. But it's not resistance to the transgenic version of the protein toxin; it's widespread resistance to Bt sprays, specifically those used to control the diamondback moth (Plutella xylostella) and the Indian meal moth (Plodia interpunctella) (8, 16). Laboratory experiments show that resistance in other insect species may follow. This article deals with the resistance issue in Bt transgenic crops in the context of Bt resistance as a whole.
 Johnny-Come-Lately
Johnny-Come-LatelyInsect resistance is old hat to entomologists. Funding for basic research toward insecticide resistance management has been sought for years, but the subject generated little interest. Now that gene jockeys can engineer pest control into plants, environmental advocacy groups (EAGs) have trotted out the pest resistance card as one of several reasons why irreversible disaster worse than DDT and nuclear power combined is at hand.
Greenpeace and others claim to fear that organic farmers will loose a valuable, natural, sustainable insect control method--Bt sprays--to the evil specter of genetically engineered Bt toxin-containing crops. The fact is that entomologists were reporting resistance to Bt sprays commonly used on organic crops nearly fifteen years ago. Waiting until now to make a fuss seems slightly opportunistic.
Bt sprays may lead to pest resistance quicker than Bt transgenic crops (12), but insects have the potential to develop resistance to both technologies. The challenge is how to deploy transgenics to slow resistance and perhaps even avert it altogether. We have learned a lot about resistance management, even though the track record with conventional pesticides is not good. But now, early in the game, is our golden opportunity to put new ideas in place for transgenics before resistance develops.
Successful management of Bt resistance will require attention to seven commonsense elements (7):
These seven elements were addressed by the registrants when the first Bt-protected crops were approved for commercial introduction in 1995. But EPA approval was conditional and up for renewal in 2001, pending results of a plethora of studies. Extensive monitoring and computer modeling are being employed to predict the evolution of resistance. Thus far, there are no reports of resistance development in field-grown transgenic crops.
Given that insects illustrate well the cliché that "you can't fool Mother Nature," will our efforts stem the evolutionary inevitability of resistance development? Let's review why insects (as well as many other organisms) become resistant in the first place, and then tackle some of the seemingly intractable obstacles and proposed solutions to successful insect resistance management.

We all know that any organism's biochemical, physiological, morphological, and behavioral characteristics ultimately depend on genes, those functional units of heredity strung like beads on chromosomes in the nucleus of every cell. The gene itself is composed of separate molecules called nucleotides that are linked together to make the DNA polymer. Any one gene is made of different numbers and combinations of the possible DNA nucleotides (also commonly called DNA bases). Every plant and animal chromosome has two helical woven strands of DNA, providing the cell with the ability to faithfully replicate its genes when it divides to form new cells. Animal cells with two copies of each chromosome (i.e., diploid), one inherited from the mother and one from the father, actually have two copies of a gene. The genetic makeup of an organism is called its genotype.
One of the DNA strands from each of the chromosomes is transcribed into an RNA polymer that eventually directs translation of the gene information into proteins. The basic function of the proteins made from each copy of the gene are the same, but the resulting characteristics or activities of the proteins translated from each gene may differ. These various copies of the gene are called alleles. All of the resulting physical characteristics or activities caused by each gene, taken collectively, are called the phenotype.
For a simplistic example, an allele of a human gene is responsible for directing synthesis of brown pigment in the eye. Humans who carry this brown allele have brown eyes. The actual gene is part of a person's genotype, but the brown pigment is part of the phenotype (or physical appearance). Individuals without the brown allele will have blue eyes because the blue allele does not direct synthesis of a pigment. If a person has one brown allele and one blue allele their eyes will be brown because the brown allele directs pigment synthesis.
The terms dominant and recessive are used when looking at the phenotype of the gene activity. In the case of human eye color, brown is dominant, because an individual with one brown and one blue allele has brown eyes. Not all gene activity can be so easily seen, but the concept of dominance of alleles is the same.
Proteins serve many varied functions in an organism: detoxification enzymes, cell receptors, biochemical messengers (such as neurotransmitters), and protein gates (structures that selectively allow ions to move into and out of membranes). Enzymes, receptors, and gates are coincidentally biochemical sites with which insecticides, including the Bt toxin proteins, are known to interact.
In any population of insects, some individuals will have copies of alleles that may produce very active detoxification enzymes that very quickly metabolize an insecticide. Other individuals may have copies of an allele producing a much less active form of the enzyme. The individual allele imparting the swift detoxification ability can be characterized as being a resistant (R) allele, whereas the less-active allele would be considered susceptible (S).
If an individual has chromosomes with only R alleles or S alleles, they are homozygous for that trait. If they have a mix of R and S alleles, they are heterozygous.
When a pest population is sprayed with an insecticide, those individuals with copies of the R alleles, especially the homozygous ones, will be more likely to survive, mate, and produce offspring that inherit the efficient detoxification alleles. Figure 1 illustrates inherited tolerance and resistance traits.
|
|
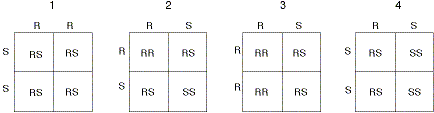 |
| Genetic characterization of offspring from matings of parents homozygous and heterozygous for resistance and susceptible genes. Each parent contributes one chromosome, which has two possible alleles, R or S (shown on the tops and sides of each box). In this case, it doesn't matter which parent has which gene pairs. Each individual square within a box represents the possible genetic makeup of the offspring, resulting from the mating. In box 1, mating of homozygous R and S parents results in all heterozygous offspring (RS). In box 2, heterozygous matings result in a 25% probability of the offspring being homozygous for RR, 25% homozygous for SS, and a 50% probability of being heterozygous. In box 3, a homozygous R and heterozygous RS mating will increase the probability of homozygous, resistant offspring to 50%. In box 4, a mating of a homozygous susceptible individual with a heterozygous individual will result in 50% probability of homozygous susceptible offspring. To reduce the speed of resistance development to Bt crops, the situations represented in boxes 1 and 4 need to be maximized. |
Receptor proteins and membrane gates also occur in different forms that may more or less readily bind with a toxin. An individual having a receptor with comparatively less penchant to bind with a toxin will likely survive exposure and pass its resistant alleles on to its offspring.
Whether the heterozygous offspring would survive an insecticide spray is dependent both upon the dose of the spray and upon which allele (R or S) exerts dominance (Figures 1 and 2). Research suggests that when the resistance mechanism is due to production of an efficient detoxification enzyme, R alleles may be semi-dominant over S alleles (13). In these situations, usually the toxin is recognized by a receptor, which in turn stimulates a cascade of events leading to death of the organism. Resistance is not conferred by an alteration of the receptor, but rather by swift detoxification of the toxin. Individuals with dominant R alleles, homozygous or heterozygous, would have the ability to detoxify the toxin but with different efficiency. Heterozygous RS individuals, having a lower concentration or activity of the detoxification enzyme than homozygous RR individuals, would succumb to a moderately lower dose of toxin. Individuals with homozygous S alleles would not have the ability to sufficiently detoxify the pesticide and would die at very low doses.
|
|
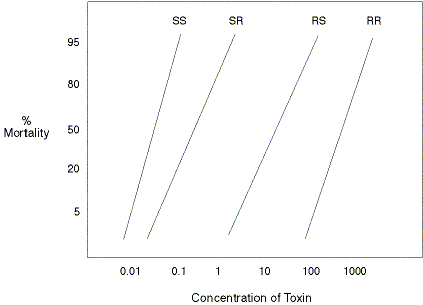 |
| Theoretical dose-response curves for insect populations with different genetic backgrounds (genotypes) for susceptibility to a toxin. The SS and RR populations contain individuals that are homozygous for susceptible and resistant genes, respectively. The SR population is heterozygous, but the S allele is semi-dominant over the R allele (i.e., the R allele is recessive). This population is killed by slightly higher concentrations of the toxin necessary to kill SS individuals. If the R allele is dominant to the S allele, the RS heterozygote will only be killed by doses significantly higher than required for the SS or SR population but less than the RR population. To delay resistance development to the Bt toxin, all the SR or RS individuals will ideally be killed. This objective will be reached more easily without having to substantially increase the toxin concentration if R is recessive. |
When the resistance mechanism is due to a change in the affinity of a receptor to bind with an insecticide, the S allele may be semi-dominant over the R allele (13). In this case, susceptible (S) insects have receptors with a strong penchant for binding to the toxin. Resistant (R) insects have receptors that bind the toxin inefficiently or perhaps not at all, and thus they do not die when exposed to normal pesticide application rates. In this scenario, the SR heterozygote would require a moderately higher dose than the SS homozygote for a lethal effect as only some of the RS receptors would recognize and bind the toxin (Figure 2).
Through natural variation, every population contains alleles that will confer tolerance to a pesticide. The speed with which the population develops resistance is influenced by many factors, among them how often the field is sprayed and the initial frequency of occurrence of the R alleles. To develop an appropriate resistance management plan for Bt crops, it's a good idea to first determine the population's natural variation for tolerance to Bt. Variation can be characterized by exposing field-collected populations to different doses of purified Bt toxin in their diet and calculating concentrations lethal to 50% and 99% of the population (i.e., the LC50 and LC99, respectively). This method is useful for finding a dose that discriminates between resistant and susceptible individuals, but it fails to inform about the frequency of resistant alleles in a population.
A second method to characterize variation calculates the frequency of resistant alleles in different pest populations. Collected insects from distinct populations are mated with known homozygous susceptible members of lab colonies and then the offspring are exposed to a discriminating dose of Bt toxin or to Bt-protected plants. The discriminating dose essentially differentiates between a homozygous susceptible individual and a heterozygous individual. The results can be used to calculate the frequency of resistant alleles. Studies of tobacco budworm (Heliothis virescens), the major pest of cotton, and the European corn borer (Ostrinia nubilalis) on corn, have estimated the pre-selection frequency of the resistant Bt toxin allele as close to one in a thousand (i.e., 10-3) (1, 5).
Entomologists have become rather fatalistic
about insecticide resistance--experience thus far tends to indicate
it will inevitably develop. But for two decades, schemes for delaying
resistance development have been proposed, and a few have been
implemented (15). The goal of such plans is to keep
resistance low enough in the field that control can be achieved
and maintained without raising application rates.
Everyone agrees that now is the time to impose insecticide resistance management (IRM) plans while the frequency of naturally Bt-resistant insects is optimally low. Hoping to keep Bt viable as a spray and a transgenic technology, EPA, USDA, industry, and academic scientists have reached consensus that the most feasible strategy to significantly delay the development of Bt resistance is to adopt the "high dose/refuge" strategy (17).
A "high dose" can be considered one that--in addition to killing all susceptible individuals--is sufficient to kill all of the potential heterozygous offspring from the mating of resistant and susceptible individuals and even many of the homozygous resistant individuals (9). With the high dose strategy, Bt crops should express enough toxin to be able to kill 100% of the susceptible insects and 99% of any of the heterozygotes. A high dose has been estimated to be about 25 times higher than the Bt toxin concentration lethal to 99% of the population (i.e., the LC99) (4, 10).
The "refuge" is an area where non-Bt crops would be planted, encouraging survival of some susceptible insects. If a sufficient population of insects with susceptible alleles survives to mate with homozygous resistant insects, the frequency of R alleles will be diluted in the next generation.
If the high dose/refuge strategy is to work, several conditions must be met.
If enough susceptible insects are present in close proximity to resistant insects, interbreeding is generally considered a sure thing. One potential problem is that all the insects must have a similar development speed and transform into adults at about the same time (known as synchronous development). Recently published research suggests that Bt-resistant pink bollworms (Pectinophora gossypiella) take longer to develop on Bt cotton than susceptible worms do on non-Bt cotton (6). Although much overlap in developmental time occurred in the experiments, enough resistant pink bollworms emerged later than the susceptible moths to cause some concern about likelihood of random mating. On the other hand, other research has shown that Bt-resistant Colorado potato beetles, which also developed more slowly on Bt potato, lost the ability to produce viable eggs (2). More research will be needed to solve the riddle of developmental time and reproductive fitness.
Ideal refuge size has been mildly controversial.
If a grower plants significant non-Bt crop acreage, he will likely
want to spray it to protect yield. If the refuge is small, however,
he may accept a possible yield loss in return for reducing insecticide
costs in the Bt crop portion of his field. Early recommendations
in cotton suggested a 4% refuge if unsprayed (7).
If a grower were going to spray the refuge, the refuge should
comprise at least 20%. (In neither case should the Bt and non-Bt
seed be mixed; each should be planted as a block in reasonable
proximity to the other.)
Genetic modeling for tobacco budworm in Bt cotton, which expresses
enough toxin to kill all heterozygotes, suggests that a non-Bt
refuge of 4% could lead to resistance development in at least
10 years (5). Thus, 4% is now seen
as insufficient. A recently published study using Bt broccoli
and diamondback moth as a model system showed that 20% unsprayed
refuges were superior to 20% sprayed refuges in minimizing proliferation
of resistant moths (14).
Based on modeling and field research in progress, factors for optimizing refuge size and structure have begun to come into focus. The USDA and EPA recently issued a joint position paper recommending refuges of specific sizes and minimum distances from the Bt crop (Table 1) (17).
|
|
||||
| Crop | Bt Gene | Refuge Size | Refuge Management | Location |
| Potato | Cry3A | 20% | Spray if needed | Next to Bt crop |
| Cotton | Cry1A(c) | 20% 40% | No spray Spray if needed | Within 1 mi. of Bt crop |
| Corn: Industry & Grower Plans | Cry1A(b) Cry1A(c) | 20% | Spray if needed | Within 0.25-0.5 mile of Bt crop |
| Corn: University Recommendations | Cry1A(b) Cry1A(c) | 20-30% 40% | No spray | Within a 320 acre section of Bt crop |
| Corn: EPA mandate | Cry9C | 20-30% | No spray | Within 1500-2000 ft of Bt crop |
| Refuge recommendations for managing development of insect resistance to Bt crops (17). "Refuge size" is the area of non-Bt crop planted as a percentage of the Bt crop area. | ||||
Determining an optimum refuge size is easier when considering a Bt crop that preferentially targets only one pest, rather than when two or more pests are involved. See the sidebar, One Glitch in the System, below, for an example of the complications in a multi-pest system.
The incredibly fast pace of technological change makes some people nervous, and Bt transgenic technology seems to have crept up on us so quickly that some felt research to prove its merit and safety was lacking. But when we go beyond the shallow newspaper and advocacy group pronouncements, we discover a significant depth of knowledge and research already in place. The technology appears to have a very low potential for human health hazards and is safe for nontarget organisms, especially insect natural enemies.
Resistance has always been a problem--some would criticize the use of Bt crops on the basis that resistance is inevitable, so why even bother. But insects can even adapt to sustainable practices like crop rotation. Witness recent research from the Corn Belt showing that corn rootworms have adapted to recommended crop rotation practices that had been responsible for cutting insecticide use in half (11) (see also related article "Western Corn Rootworms Adapt to Crop Rotation" in April 1999 AENews No. 156). Should growers not have adapted corn-soybean rotations because one day rootworms would outsmart them?
Years of thinking about IRM have laid the groundwork for the opportunity to manage resistance against Bt crops from the start and achieve long-term, successful use of one more crop protection tool. Many scientists are in favor of this technology because it can cut synthetic pesticide use, protect biocontrol options, and augment yield without having to use more land for farming.
Genetics-based population models indicate that IRM should work if everyone follows the guidelines. EPA has mandated IRM plans since the introduction of Bt crops. Continued monitoring of changes in pest susceptibility is a requirement of the IRM programs. Perhaps, for the first time, there is widespread agreement among all stakeholders of what needs to be done. I hope that policy makers will resist the popular urgings to condemn genetic engineering technologies wholesale. As the National Academy of Science recommended in its recently released report, we must judge the pest-protected plant technology on a case-by-case basis (10).
|
The Corn Earworm/Cotton Bollworm Problem When considering a Bt crop that preferentially targets only one pest, determining an optimum refuge size is easier than when two or more pests are involved. A problem occurs when the secondary pests are not as susceptible to the Bt toxin as the primary pest. Such is the case for the common secondary pest of corn and cotton, Helicopvera zea, known variously as the corn earworm and cotton bollworm. Bt cotton is very effective against the primary cotton pest, the tobacco budworm, but it confers, at most, 90% mortality in the secondary pest, the cotton bollworm. Computer modeling shows that this comparatively lower mortality could cause resistance to develop among cotton bollworms in three to four years (5), suggesting the need for a much bigger refuge area. Similarly, the corn earworm is less susceptible than the European corn borer to the registered Bt toxins. Comparatively fewer kills of earworms will result in more rapid resistance development. The real problem comes in the South, where earworms first invade corn and then move into cotton (as the bollworm). As if this situation isn't bad enough, the earworm/bollworm has many alternate crop hosts besides corn and cotton, so resistance development to Bt could have the potential to affect control by Bt sprays. On the other hand, some commercial Bt sprays have several different toxin proteins in them that might help overcome resistance to one specific toxin. Nevertheless, to alleviate the possibility that earworms will develop resistance, EPA has mandated the following IRM (17). Bt corn based on the Cry1A(c) gene cannot be planted in cotton regions because Bt cotton also contains the same gene. If Bt corn contains the Cry1A(b) gene resulting from the genetic engineering events* known as MON810, BT11, and DBT418, then a 50% refuge must be planted in fields within 0.25-0.5 miles of a cotton crop. These latter Bt corn plants express the toxin in the silks and kernels, which are food sources for the earworms. No special refuges are required for Bt corn based on engineering event 176; the Cry1A(b) toxin is not expressed in the silks and kernels. The Bt line based on Cry9C is not toxic to the earworm, so no special refuges will be required because there will be no resistance selection pressure. *ED. NOTE: Every laboratory insertion of the Bt gene is called an "event." These events are tracked and numbered. While the gene Cry1A(b) is the same in various engineering events, the resulting expression among the plants' tissues varies from one event to another. |
Dr. Allan S. Felsot is an Environmental Toxicologist with the Food and Environmental Quality Laboratory at WSU. He can be reached at (509) 372-7365 or afelsot@tricity.wsu.edu.
In the April
issue of Agrichemical and Environmental News (AENews No. 168),
I wrote about the special
chemistry involved with moth sex pheromones. This month I
will address the biological aspects of the attractant process.
The insect order Lepidoptera (moths and butterflies) is one of nine orders of insects that undergo complete metamorphosis. Larvae or juveniles of such orders bear little resemblance to their adult parents. Accordingly, they typically exploit a very different food source than their adult counterparts and occupy an entirely different ecological niche. In the case of moths, the first morph (larval stage) is devoted to the arduous labor of nutrition and growth, while the adult morph is the time for reproduction.
Most female moths are selective in where they choose to lay their eggs. For almost all moth species, parental participation and care end with site selection; the adults move on. After hatching, larvae or caterpillars are on their own, and they have one very basic task: eat and eat and eat!
Moth larvae eat and grow their way through five instars (physical states on their way to adulthood). Physical structure during this period is relatively simple: primarily mouth, gut, and anus. Reproductive organs remain in an embryonic state. Each larval instar is separated by a molt (skin shedding).
When the fifth instar stage is complete, the larva molts and forms a pupa, which externally appears quiescent but internally is extremely metabolically active. During this time, fat accumulated during larval growth is converted to adult tissues, including reproductive organs, wings, and antennae.
Lepidoptera live as individuals, dispersed throughout their territory, as opposed to being tightly grouped in a colony. This presents obvious advantages when in the larval stage: greater food resources are available for each individual and the population's risk of annihilation from predation or parasitism is reduced. But with the eclosion (emergence) of the adult morph from the pupa, the moth's focus changes. While larvae are eating machines, moths are sexual beings-the early part of their adult lives is devoted to finding a mate.
So, what's a lonely moth to do? Moths have solved the problem of geography: females excrete attractant pheromones and males are irrepressibly drawn by these attractants.
The female sex pheromone glands of tortricids (e.g., codling moth, oriental fruit moth, fruit tree leafroller, red-headed fireworm, Mexican jumping bean moth) are located dorsally (on the back side) between the eighth and terminal abdominal segments. Development of the pheromone gland in the redbanded leafroller moth has been investigated thoroughly by Feng and Roelofs (1). They found that reorganization of the caudal (rear) end of the individual is completed within sixty hours after the formation of the pupa. Differentiation of the pheromone gland begins about twelve hours later. From about four days after pupation begins until the emergence of the adult from the pupa (a total of about six and a half days from the beginning of pupation), the glands grow. Pheromone production begins a day before eclosion and peaks at three to four days after eclosion, reaching about 80 nanograms per day at peak.

Pheromone detectors are found only on the antennae of males. Male silkworm moths, for example, have large feathery antennae that are studded with about 17,000 sense hairs devoted exclusively to sensing pheromones (3). The walls of each hair have thousands of tiny pores. Tubules connect the pores to the hair's fluid-filled center, which contains nerve endings (dendrites). Once a pheromone molecule is absorbed on the antenna, it diffuses into a pore and through a tubule to chemoreceptors on the dendrite. The receptors conform to the molecular shape of a pheromone component unique to that species in a lock-and-key fashion (see AENews 168 "The Chemistry of Moth Pheromones"). When the pheromone "key" find's the corresponding male's receptor "lock," a neuron is stimulated to fire, sending a signal to the brain.
The antenna of a male silk moth is a very efficient sieve for a low concentration of molecules. Each sense hair can comprise hundreds of sensilla. When as few as 200 sensilla send a signal to the moth's brain, an anemotaxic response occurs-the male reorients his flight into the wind. Once closer to the source-point female, the male moth demonstrates a chemotaxic response-moving toward a greater concentration of the pheromone. In close proximity to pheromone-emitting females, the sensilla on the antennae of male moths send strong signals to their brains, and courtship behaviors begin. If the female deems the male worthy-i.e., sufficiently athletic and energetic in his courtship behaviors-copulation occurs.
Most female moths are monandrous: they mate only one time with a single male. Accordingly, mated females of these species stop sending out strong pheromone signals. Tang et al. observed that pheromone release by female gypsy moths falls below detectable levels within twenty-four hours following successful copulation (4).
Multiple matings of tortricids have been observed, however (2). Male lightbrown apple moths (Epiphyas postvittana) are capable of mating on average 6.6 times. Multiple mating of a female is related to the potency of the male with which she initially mates, as chemically determined by the female after copulation. Foster and Ayers observed that about 20% of virgin females that mated with a virgin male chose to mate again (2). Virgin females that mated with a previously mated male were more likely to remate, and that likelihood increased with the number of previous matings of the male. The male's mating history also affected subsequent pheromone production in the female. Females mating with virgin males exhibited a greater reduction in pheromone production than those mating with previously mated males.
Expanded knowledge regarding moth pheromone chemistry and biology has led pheromone chemists and applied biologists to great advances in the application of pheromone technologies to agroecosystems. In my next article, I will detail how moth pheromones have been effectively commercialized for monitoring and control of several important pest species.
Dr. Douglas B. Walsh is an Agrichemical and Environmental Education Specialist with WSU's Food and Environmental Quality Laboratory. His office is at the Irrigated Agricultural Research and Extension Center (IAREC) in Prosser, and he can be reached at dwalsh@tricity.wsu.edu or (509) 786-2226.
Return to Table of Contents for the June 2000 issue
Andrew Thomson, CSANR Technical Assistant, WSU
"Sustainable agriculture" has been defined as a system "that equitably balances concerns of environmental soundness, economic viability, and social justice among all sectors of society" (2). Scores of other definitions have been advanced, many with intrinsic incongruities (3). While a firm definition achieved by consensus may be important when considering policy formulation, it may not be critical at the practical level of implementation. Indeed, a flexible concept that addresses the three often-recognized components of economic, environmental, and social concern may provide a working definition sufficient for practical application.
This article looks at a case study on Lopez Island, one of the islands in Washington's San Juan County.
Lopez Island agriculture is characterized by small farms, with a diverse group of farmers including those who choose to maintain conventional farming methodology and those who have adopted alternative means of agricultural production. On Lopez, as elsewhere, sustainable agriculture can be a part of either conventional or alternative methodology. Conventional producers can incorporate more sustainable practices into existing operations, while alternative operations (including those that might be referred to as "organic," "biodynamic," or "permacultural") might seek a completely new way of operating that is sustainable into the indefinite future.
Approximately forty active
farms on Lopez Island produce products for sale to the public.
About half claim to be farming conventionally, about forty percent
claim to be farming alternatively, and about ten percent use components
from both.
 Though Lopez, like the other islands
in San Juan County, is not a major contributor to the total food
balance of the state or nation, it can serve as an illustration
of a certain type of agroecosystem within a certain cultural setting.
A 1989 public opinion survey found that Lopez residents put a
high priority on preserving agricultural land, the natural environment,
and a rural lifestyle (5). Further, they felt that a local food system would
provide food security on the island.
Though Lopez, like the other islands
in San Juan County, is not a major contributor to the total food
balance of the state or nation, it can serve as an illustration
of a certain type of agroecosystem within a certain cultural setting.
A 1989 public opinion survey found that Lopez residents put a
high priority on preserving agricultural land, the natural environment,
and a rural lifestyle (5). Further, they felt that a local food system would
provide food security on the island.
In what ways and to what extent have alternative agricultural approaches proven beneficial to the Lopez community? A three-month observation period and a survey were completed during the past year. Some of the findings can be organized under the three broad areas most often conceptualized as components of sustainable agriculture: economic, environmental, and social.
As the editor of Farm Journal wrote, "What is sustainable agriculture, after all? The only sustainable agriculture is profitable agriculture" (1). Certainly, if agriculture is not profitable, something must change.
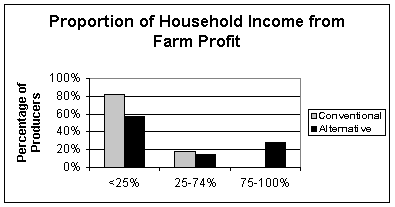
For the majority (seventy-one percent) of Lopez Island farmers, farming is a part-time endeavor; many hold second jobs. This makes it is very difficult to measure agricultural economic sustainability because agricultural income is not the only source of income in many Lopez agricultural households. Nevertheless, results from the survey show that, for nearly thirty percent of self-proclaimed "alternative" farmers on Lopez Island, farm profits contribute seventy-five to one hundred percent of their household income. Fifty-seven percent of the alternative growers derive less than twenty-five percent of their income from their farm. In contrast, none of the self-proclaimed "conventional" farmers reported receiving seventy-five or more percent of their household income from their farm, and eighty-two percent derived less than twenty-five percent (Figure 1). This is not to say that alternative producers are making more money. In fact, results show that there is not a significant difference between alternative and conventional producers' total net earnings. But economic sustainability cannot be measured by cash earnings alone. A producer's non-cash contribution to the household (for example, food that is provided from the farm) must be considered, as well as lifestyle differences and perceived income needs. For this reason, proportion of household income may be a better measure of economic sustainability.
|
|
 |
|
|
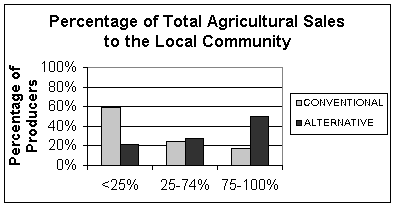
Selling habits of alternative versus conventional producers on Lopez Island may account for part of the proportion-of-household-income discrepancy. About half of the alternative producers sell three quarters or more of their products locally, compared to only eighteen percent of conventional producers (Figure 2). Local product sales reduce transportation costs and eliminate middlemen. (Lopez Island's primary link to mainland Washington is the state ferry system, a relatively expensive mode of transport.) In addition, many alternative farmers are producing organic products, which can fetch a higher return and may increase profitability for the producer.
|
|
 |
|
|
Another key component of economic sustainability on the island is the county's Open Space Agriculture Program, which provides farmers with a significant reduction in property taxes if land is kept in agricultural production (5). As property values continue to escalate in the area, this program provides incentive for farmers to continue farming and for purchasers of previous or potential agricultural land to continue or begin farming.
The Lopez Island ecosystem is fragile; it closely approximates a closed system. There are no rivers to carry away contaminants; what goes into the groundwater will eventually surface again locally. There is little migration; wildlife species remain on the island through many generations. This kind of sensitive environment needs careful attention to maintain its natural balance.
Most of the alternative producers on Lopez Island use minimal or no synthetic pesticides or fertilizers. With approximately forty percent of producers on Lopez claiming to farm alternatively, substantially less than the normal amount of synthetic chemicals are used. While some may argue that synthetic pesticides and fertilizers cause little harm to the environment, the fact remains that a lower level of pesticide use reduces the potential for controversy on this score.
The community on Lopez Island has shown a strong commitment to sustainable agriculture. One of the driving forces behind this effort is the Lopez Community Land Trust (LCLT), an organization with goals to provide affordable housing on Lopez Island as well as build food and economic security through Sustainable Agriculture and Rural Development (SARD) and Community Based Economic Development (CBED) programs. About half of the island's alternative producers and nearly a fifth of the conventional producers are involved with the SARD component of the LCLT program. Several successful programs supporting sustainable agriculture concepts have been initiated by the LCLT.
- The Lopez Island Farm Products Guide, published annually, tells island consumers where and how to purchase local farmers' products.
- LCLT provides financial support for a "hands-on" internship program established in 1990. Interns come from universities to learn about sustainable agriculture and rural development by working fifteen hours per week with local farmers and fifteen additional hours per week at the Land Trust office in exchange for college credit and a weekly stipend.
- The Learning Food Garden is a program that gives local middle school students the opportunity to learn about growing and selling organic produce while providing them with school credit as they participate in an actual farming operation through an entire season. The LCLT provides the farmer/teacher and students with tools, seed, a small stipend, and a campout at the end of the season.
- Seed Savers was established to develop a self-sufficient local seed bank. Participants are responsible for producing seed from a particular vegetable, which the seed bank in turn makes available to local producers and the public.
- The LCLT is in the process of developing a local USDA-licensed food-processing center for use by San Juan County producers. To be known as the Lopez Community Food Processing Center (FPC), its goal is decreasing county dependence on imported food products by providing facilities, services, and education that support a stronger community-based food system. The FPC will provide processing facilities for products grown in San Juan County for small-scale family and commercial processing (4).
Another measure of the social aspect of sustainability is the amount of labor required by subject farmers. The survey indicated that half of alternative farms and about a fifth of conventional farms rely on seasonal to year-round employees. Most of this work is performed by community members, who are often paid or otherwise compensated. Working together in this fashion fosters community interaction, a key component of community development.
Sustainability in agricultural systems is viewed by many as a desirable goal. In practice, attaining sustainability can prove challenging. It is not necessary that every farm be completely sustainable to be considered successful. Farmers and communities achieve a victory by attempting to incorporate as many sustainable agricultural practices into their system as are feasible under their particular set of environmental conditions and circumstances. As the Lopez Island case study has shown, selecting alternatives that work well within a particular set of circumstances may help to improve the economic, environmental, and social sustainability of agriculture within an individual region. Regions vary, conditions and knowledge are constantly changing, and growers can and must adapt their practices to address current and future realities. There will be no "one-size-fits-all" solution for sustainable agriculture. What must be held constant is sustainable thinking.
Andrew Thomson is Technical Assistant for the Center for Sustaining Agriculture and Natural Resources (CSANR) at Washington State University. He can be reached at athomson@wsu.edu or (509) 338-0879.
Return to Table of Contents for the June 2000 issue
|
The Pesticide Information Center (PIC) at Washington State University Tri-Cities maintains a searchable database of information contained in the pesticide labels registered in Oregon and Washington states. PIC staff continually updates and improves the Pesticide Information Center On-Line (affectionately known as "PICOL") label database. Recently we made two such improvements. First, some helpful (we hope) information has been added to the crop dictionary section of the database: a new field containing notes defining the crop codes and referring users to other related codes. For example, if you enter the term "corn" into the crop dictionary search field, you will see listings that remind you to also search using popcorn, sweet corn, field corn, silage, corn seed, and possibly stored grain, depending upon your interest. If you take a look at the entries for grass you will see a note defining this term specifically as grasses grown for agricultural use; a notation that lawn and turf are separately defined crop terms; and listings of other related crop codes. The second improvement concerns adjuvants (for which registration is required in Washington State). We are now coding the adjuvants into six separate "type" categories: antifoaming agents, buffers, drift control agents, surfactants, extender/stickers, and those adjuvants containing ammonium sulfate. (An "Adjuvents: Other" category exists for those with ill-defined functions or functions not covered by the six main categories.) If an adjuvant makes claims to serve several functions, it will be assigned multiple codes. We hope these changes will make the database not only more useful but also more straightforward. Should you have any questions or comments about the PICOL label database, please contact Jane Thomas at (509) 372-7493 or jmthomas@tricity.wsu.edu. |
Return to Table of Contents for the June 2000 issue
Washington Pest Consultants Association organizes an annual series of collection dates and sites for empty pesticide containers. Dates and locations are subject to change; it may be wise to confirm with a telephone call before participating. Contact telephone numbers for specific events are given in the table below. For general questions, or if you are interested in hosting an event at your farm, business, or in a central location in your area, contact Northwest Ag Plastics representative Clarke Brown at (509) 965-6809 or David Brown at (509) 469-2550 or dbrownwash@msn.com. More information on pesticide waste and container recycling is available on the WSU Pesticide and Environmental Stewardship site.
| June 1 | 8a-10a | Davenport Airport | Northwest Aviation Inc | Lee Swain | (509) 725-0011 |
| 1p-3p | Harrington | Western Farm Service | Jim Baye | (509) 253-4311 | |
| June 2 | 8a-10a | Ritzville | Cenex Grange Supply | Dale Anderson | (509) 659-1360 |
| 1p-3p | Odessa | Cenex Grange Supply | Greg Luiten | (509) 982-2693 | |
| 4p-6p | Lind | Rudy's Aviation | Rudy Fichtenberg | (509) 677-8858 | |
| June 5 | 8a-10a | Mattawa | Wilbur Ellis | Al Hilliker | (509) 932-4988 |
| 11a-Noon | Royal City | Saddle Mountain | Mike Pack | (509) 346-2291 | |
| 2p-5p | Royal City | Cenex | Ted Freeman | (509) 346-2213 | |
| June 6 | 8a-1p | Othello Airport | Conner Flying Inc. | Mark Conner | (509) 488-2921 |
| 1p-2p | Othello | B&H Chemical | Larry Hawley | (509) 488-6576 | |
| 3p-5p | Bruce | Cenex | Lori Anderson | (509) 488-5261 | |
| June 7 | 8a-11a | Moses Lake | Tom Dent Aviation | Tom Dent | (509) 765-6926 |
| 2p-5p | Warden | D&S Crop Care | David Smith | (509) 349-7660 | |
| June 8 | 1p-3p | Almira | Cenex | Don Felker | (509) 639-2421 |
| June 9 | 8a-11a | Wenatchee | Fieldmen's Assoc. | Floyd Stutzman | (509) 669-0420 |
| June 13 | 8a-Noon | Prosser | Simplot | John Cullen | (509) 837-6261 |
| 2p-5p | Sunnyside | Bleyhl Farm Service | Gary Herdon | (509) 839-4200 | |
| June 14 | 8a-Noon | Zillah | Bleyhl Farm Service | Ray Oversby | (509) 829-6922 |
| June 19 | 8a-10a | Spokane | WSDA | Tim Schultz | (509) 533-2690 |
| 222 N Havana | WSU | Jim Lindstrom | (509) 533-2686 | ||
| 11a-1p | Mead | Cenex | Todd Race | (509) 466-5192 | |
| 3p-5p | Deer Park | Inland Agronomy | Jim McAdam | (509) 276-2611 | |
| June 20 | 8a-9a | Fairfield | Wilbur Ellis | Ric Murison | (509) 283-2411 |
| 10a-11a | Waverly | Wilbur Ellis | Monte Bareither | (509) 283-2432 | |
| 1p-3p | Tekoa | McGregor's | Charles Wedin | (509) 284-5391 | |
| June 21 | 8a-10a | Oakesdale | Wilbur Ellis | Jerry Jeske | (509) 245-4511 |
| 11a-1p | Rosalia | Western Farm Service | John Hartley | (509) 523-6811 | |
| 3p-5p | St John | McGregor's | Rick Bafus | (509) 648-3218 | |
| June 22 | 8a-Noon | Colfax | Grange Supply | Darrel Tyler | (509) 397-4353 |
| June 23 | 8a-10a | Pullman | McGregor's | Larry Schlenker | (509) 332-2551 |
| 11a-1p | Palouse | Dale's Flying Service | Dale Schoeflin | (509) 878-1531 | |
| June 30 | 10a-Noon | Quincy | Cobia Spray Service | Jim Cobia | (509) 785-6771 |
| 1p-4p | Warden | Wilbur Ellis | Norman Parton | (509) 349-2333 | |
| July 3 | 8p-11p | Harrah | Ag Air | Lenard Beierle | (509) 865-1970 |
| July 10 | 8a-Noon | Walla Walla | McGregor's | Gary Burt | (509) 529-6787 |
| 1p-3p | Waitsburg | McGregor's | Terry Jacoy | (509) 337-6621 | |
| July 12 | 8a-10a | Prescott | Agri Northwest | Shawn Elder | (509) 547-8870 |
| 11a-2p | Prescott | Broetje's Orchard | Joe Shelton | (509) 749-2217 | |
| Prescott | Flat Top Ranch | Dave Hovde | (509) 547-9682 | ||
| 3p-5p | Pasco | Air Trac | Gerald Titus | (509) 547-5301 | |
| July 13 | 8a-11a | Eltopia | Wilbur Ellis | Vern Record | (509) 297-4291 |
| 1p-3p | Eltopia | Eastern Wa Spray Serv. | Willis Maxon | (509) 297-4387 | |
| July 14 | 8a-Noon | Connell | B&R Crop Care | Chris Eskildsen | (509) 234-7791 |
| 1p-3p | Pasco | Pfister Crop Care | Steve Pfister | (509) 297-4304 | |
| July 17 | 8a-10a | Oroville | Northwest Wholesale | Herb Teas | (509) 662-2141 |
| 11a-3p | Tonasket | Wilbur Ellis | Brian Hendricks | (509) 682-5315 | |
| July 18 | 8a-10a | Okanogan | Okanogan Air Service | Bill Lockwood | (509) 422-2617 |
| 1p-4p | Brewster | Wilbur Ellis | Brian Hendricks | (509) 682-5315 | |
| July 19 | 8a-Noon | Chelan | Wilbur Ellis | Brian Hendricks | (509) 682-5315 |
| July 20 | 8a-Noon | Yakima | Wilbur Ellis | Doug Whitner | (509) 248-6171 |
| July 31 | 8a-11a | Wilbur Airport | Greg's Crop Care | Greg Leyva | (509) 647-2441 |
| 2p-4p | Davenport Airport | Northwest Aviation Inc | Lee Swain | (509) 725-0011 | |
| Aug. 1 | 8a-11a | St John | McGregor's | Rick Bafus | (509) 648-3218 |
| 2p-4p | Mockonema | McGregor's | Dale Deerkop | (509) 635-1591 | |
| Aug. 2 | 8a-11a | Garfield | Cascade Flying Service | Doran Rogers | (509) 635-1212 |
| 1p-4p | Palouse | Dale's Flying Service | Dale Schoeflin | (509) 878-1531 | |
| Aug. 3 | 8a-10a | Pullman | McGregor's | Larry Schlenker | (509) 332-2551 |
| 1p-3p | Dusty | Dusty Farm Co-Op Inc. | John Stoner | (509) 397-3111 | |
| Aug. 4 | 8a-11a | Clarkston | Valley Helo Service | James D. Pope | (509) 758-1900 |
| 1p-3p | Pomeroy | McGregor's | Mark Welter | (509) 843-1468 | |
| Aug. 8 | 8a-11a | Dayton | McGregor's | Doug Wendt | (509) 382-4704 |
| 1p-4p | Waitsburg | McGregor's | Terry Jacoy | (509) 337-6621 | |
| Aug. 9 | 8a-11a | Eltopia | Wilbur Ellis | Vern Record | (509) 297-4304 |
| 1p-3p | Pasco | Pfister Crop Care | Steve Pfister | (509) 297-4304 | |
| 4p-5p | Pasco, Kahlotus Rd. | Air Trac | Gerald Titus | (509) 547-5301 | |
| Aug. 10 | 8a-10a | Eltopia | Eastern Wa Spray Service | Willis Maxson | (509) 297-4387 |
| 11a-2p | Connell | B&R Crop Care | Chris Eskildsen | (509) 234-7791 | |
| 4p-6p | Othello | B&H Ag Chemical | Larry Hawley | (509) 488-6576 | |
| Aug. 11 | 8a-11a | Othello Airport | Conner Flying Inc. | Mark Conner | (509) 488-2921 |
| 2p-5p | Moses Lake | Moses Lake Air Service | Perry Davis | (509) 765-7689 | |
| Aug. 14 | 8a-11a | Quincy | Wilbur Ellis | Dale Martin | (509) 787-4433 |
| 1p-3p | Quincy | Quincy Flying Service | Richard Weaver | (509) 787-3223 | |
| Aug. 15 | 8a-10a | Royal City | Cenex | Ted Freeman | (509) 346-2213 |
| 11a-1p | Royal City | Saddle Mountain | Mike Pack | (509) 346-2291 | |
| 2p-5p | Mattawa | Wilbur Ellis | Al Hilliker | (509) 932-4988 | |
| Aug. 16 | 8a-11a | Ephrata | The Crop Duster | Martin Shaw | (509) 754-3461 |
| 1p-3p | White Trail | The Crop Duster | Martin Shaw | (509) 754-3461 | |
| Aug. 21 | 8a-11a | Toppenish | Western Farm Service | Steve Laws | (509) 865-2045 |
| 1p-4p | Harrah | Husch & Husch | Allen Husch | (509) 848-2951 | |
| Aug. 22 | 8a-11a | Cowiche | D&M Chemical | Dee Gargus | (509) 678-5750 |
| Sept. 5 | 8a-11a | Chelan | Northwest Wholesale | Herb Teas | (509) 662-2141 |
| Sept. 6 | 8a-11a | WenatcheeTree Fruit Stn. | Fieldmen's Assoc. | Floyd Stutzman | (509) 669-0420 |
| Sept. 11 | 9a-11a | St John | Gossard Aviation Inc. | Wesley Gossard | (509) 648-3722 |
| 1p-3p | Pine City | Reed Aviation | Pete Reed | (509) 523-3950 | |
| Sept. 12 | 8a-10a | Warden | Kilmer Crop Dusting | Terry Kilmer | (509) 349-2491 |
| 11a-1p | Bruce | Simplot | Chuck Spytex | (509) 488-2132 | |
| 3p-5p | Othello | South Saddle Orchard | Mike Macy | (509) 539-5836 | |
| Sept. 14 | 8a-11a | Zillah | Bleyhl Farm Service | Ray Oversby | (509) 829-6922 |
| Oct. 3 | 9a-10a | Ellensburg | DOT | Susanne Tarr | (509) 962-7577 |
| Oct. 13 | 8a-3p | Othello | Conner Flying Inc. | Mark Conner | (509) 488-2921 |
| Oct. 16 | 8a-10a | Waterville | Western Farm Service | Dale Gromley | (509) 745-8857 |
| 11a-2p | Coulee City | Western Farm Service | Pete Thiry | (509) 632-5697 | |
| 3p-5p | Ephrata | The Crop Duster | Martin Shaw | (509) 754-3461 | |
| Oct. 17 | 8a-11a | Wilbur Airport | Greg's Crop Care | Greg Leyva | (509) 647-2441 |
| 1p-4p | Davenport Airport | Northwest Aviation Inc. | Lee Swain | (509) 725-0011 | |
| Oct. 18 | 8a-Noon | Rosalia | Western Farm Service | John Hartley | (509) 523-6811 |
| 1p-3p | Mockonema | McGregor's | Dale Deerkop | (509) 635-1591 | |
| Oct. 19 | 8a-10a | Connell | B&R Aerial Crop Care | Chris Eskildsen | (509) 234-7791 |
| 1p-3p | Pasco, Kahlotus Rd. | Air Trac | Gerald Titus | (509) 547-5301 | |
| Oct. 23 | 8a-10a | Spokane | WSDA | Tim Schultz | (509) 533-2690 |
| 222 N Havana | WSU | Jim Lindstrom | (509) 533-2686 | ||
| 11a-1p | Mead | Cenex | Todd Race | (509) 466-5192 | |
| 3p-5p | Deer Park | Inland Agronomy | Jim McAdams | (509) 276-2611 | |
| Oct. 26 | 9a-Noon | Coulee City | Cenex | Huck Dilling | (509) 632-5292 |
| 1p-3p | Almira | Cenex | Don Felker | (509) 639-2421 | |
| Oct. 27 | 8a-Noon | Moses Lake | Tom Dent Aviation | Tom Dent | (509) 765-6926 |
| 2p-5p | Warden | Kilmer Crop Dusting | Terry Kilmer | (509) 349-2491 | |
| Oct. 30 | 9a-3p | Outlook | Snipes Mtn. Trans. Stn. | Mark Nedrow | (509) 574-2472 |
| Oct. 31 | 8:30a-3p | Yakima | Terrace Hts. Landfill | Mark Nedrow | (509) 574-2472 |
Return to Table of Contents for the June 2000 issue
In reviewing the April postings in the Federal Register, we found the following items that may be of interest to the readers of Agrichemical and Environmental News.
In the April 19 Federal Register, EPA issued an order finalizing the use deletions and product cancellations for azinphos methyl that had previously been agreed upon with the registrants and that were announced in the December 3, 1999, Federal Register. Azinphos methyl products are now only permitted to be distributed or sold in accordance with the existing stocks provisions set forth in this same notice. (Page 20966)
In the April 26 Federal Register, EPA announced that the revised risk assessment for coumaphos was available for review and comment. An electronic copy of this document is available at http://www.epa.gov/pesticides/op/coumaphos.htm. Comments on this risk assessment must be submitted to EPA on or before June 26, 2000. (Page 24468)
FQPA requires that EPA periodically review pesticide registrations to ensure that, over time, they continue to meet statutory standards for safety. This review is required on a 15-year cycle. In the April 26, Federal Register, EPA issued an advance notice of proposed rulemaking (ANPRM) to alert stakeholders that EPA is beginning development of procedural regulations for this registration review. In the notice EPA: explains its preliminary interpretation of the authorizing legislation, presents goals in implementing the statutory provisions, presents the initial concept of how the registration review program might operate, identifies several issues that should be addressed in developing the program, and invites public comment on these and other issues relating to registration review. Comments on this ANPRM must be received by EPA on or before June 26, 2000. (Page 24585)
In the April 28 Federal Register, EPA announced that the revised risk assessments for chlorpyrifos-methyl and trichlorfon were available for review and comment. Electronic copies of these documents are available at http://www.epa.gov/pesticides/op/chlorpyrifos-methyl.htm and http://www.epa.gov/pesticides/op/trichlorfon.htm. Comments on these risk assessments must be submitted to EPA on or before June 27, 2000. (Page 24954)
The PNN is operated by WSU's Pesticide Information Center for the Washington State Commission on Pesticide Registration. The system is designed to distribute pesticide registration and label change information to groups representing Washington's pesticide users. PNN notifications are now available on-line. To review those sent out in the month two months prior to this issue's date, either access the PNN page via the Pesticide Information Center On-Line (PICOL) Main Page on URL http://picol.cahe.wsu.edu/ or directly via URL http://www.tricity.wsu.edu/~mantone/pl-newpnn.html. We hope that this new electronic format will be useful. Please let us know what you think by submitting comments via e-mail to Jane Thomas at jmthomas@tricity.wsu.edu.
| Chemical (type) | Federal Register | Tolerance (ppm) | Commodity (raw) |
|
||
|
|
|
|
||||
| spinosad (insecticide) | 4/5/00 page 17773 | 0.2 | apple | No | N/A | N/A |
| 0.02 | animal feed, nongrass group | |||||
| 0.02 | barley, grain | |||||
| 0.02 | teosinte, grain | |||||
| Comment: These values are corrections to tolerances that were published in the January 12 Federal Register. | ||||||
| fenhexamid (fungicide) | 4/13/00 page 19842 | 1 | prune, dried | No | N/A | N/A |
| 0.05 | plum (fresh prune) | |||||
| 6 | stone fruit, except plum (fresh prune) | |||||
| thiabendazol (fungicide) | 4/26/00 page 24398 | 0.1 | lentils | Yes | Extension | 12/31/01 |
| Comment: This time-limited tolerance is being extended in response to repeat Section 18 requests from Idaho, Washington, North Dakota, and Montana for the use of thiabendazol to control of ascochyta blight on lentils for this year's growing season. | ||||||
| fenpropathrin (insecticide) | 4/26/00 page 24392 | 0.5 | squash/cucumber subgroup | No | N/A | N/A |
W.S.U. Pullman Home Page Comments and questions: cdaniels@tricity.wsu.edu Technical Assistance: scoates@tricity.wsu.edu Copyright © Washington State University / Disclaimer Electronic Publishing and Appropriate Use Policy University Information: 509/335-3564